
This blog post is provided by David Outomuro and Alberto Corral-Lopez and tells their #StoryBehindthePaper for the article “Field evidence for colour mimicry overshadowing morphological mimicry”, which was recently published in the Journal of Animal Ecology.
Intuitively, to gain higher protection from predators, mimic species should evolve to be mirror images of their models. However, mimic species with very poor resemblance to their models are commonplace in nature. This so-called imperfect mimicry has indeed been puzzling for biologists for a long time. Previous laboratory experiments have demonstrated that one of the causes behind this phenomenon is related to predator perception biases. Predators can pay more attention to certain prey signals than others. For example, birds are known to pay more attention to colours over patterns or shapes. Consequently, patterns and shapes would be expected to be more poorly resembled than colours, leading to the observation of imperfect mimicry. Gathering evidence for this phenomenon outside laboratory settings has been challenging. In our article published in the Journal of Animal Ecology, we present one of the first studies investigating predator perception biases in wild populations.
The focus of our study was the “clear wing complex” mimicry ring (a group of species within a local community sharing an aposematic signal). Mainly distributed in tropical areas of Central and South America, this mimicry ring is dominated by glasswing butterflies (Fig. 1). The UV-reflective white patch that these butterflies present on their mostly transparent wings functions as an honest signal of their unpalatability, which originated from accumulated alkaloids. While a white wing patch that reflects ultraviolet is not a common warning signal found in nature, it seems to effectively advertise the message to their main predators (birds) in this habitat. Indeed, many insect species have evolved a similar aposematic signal mimicking glasswing butterflies. Interestingly, among these mimic species, one can find the only family of damselflies known to date to have evolved mimicry, the family Polythoridae (Fig. 1).

In a previous published study in this system, we characterized the complexity of the aposematic signal between glasswing butterflies and Polythoridae damselflies, and found that they share similarities in multiple characteristics, including wing colour, wing morphology and flight style (DOI: 10.1111/evo.12963). New questions were raised with such findings: How do bird predators perceive the differences in their warning signals? How are damselflies producing similar signals to those present in glasswing butterflies? Olof Leimar’s group at the University of Stockholm had recently published an influential series of studies on how stimulus salience can explain the evolution of imperfect mimicry, particularly using bird predators (e.g. DOI: 10.1016/j.cub.2014.02.061). How could we test for stimulus salience in our particular system? We got the answer to this question during the Evolution meeting in Austin, Texas, where we both attended an interesting talk by Dr. Susan Finkbeiner on the use of artificial replicas of butterflies in the field to study mimicry evolution (e.g. DOI: 10.1111/evo.13165).
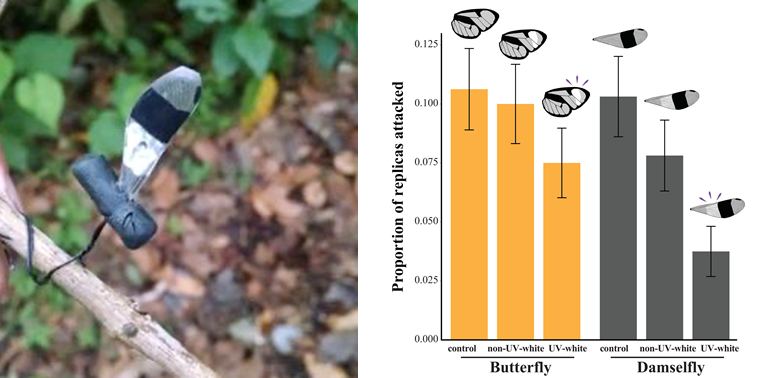
Next, the most exciting part of the study took place, the fieldwork. Well, probably after convincing the panel of two Foundations to grant us the money to run a field experiment testing predation on replicas of glasswing butterflies and damselflies in Colombia. And probably also after manually putting together the 2000 artificial replicas before flying there (Fig.2). We made replicas that looked like butterflies or damselflies perching, and we also added a colouration treatment: without the white wing patch, with an UV-reflective white wing patch, or with a non-UV-reflective white wing patch. Eventually, we travelled to Bogotá, where we joined our collaborators from Universidad de los Andes.

We then headed to the piedmont habitat (the transition between the cloud and the rain forest on the western slopes of the western Andes mountain range) (Fig. 3). There, we ran the predation experiment for an intense period of 5 days, hiking close to 20 km daily to place our artificial replicas and evaluate predator responses towards them.

We found that predator attacks on artificial replicas differed between the colouration treatments, but not between the species, suggesting that colouration is more important than morphology, i.e. colouration overshadows morphology for bird predators (Fig. 4).
We additionally explored the extent of the aposematic colour in the local community of insects, and how so different insects as butterflies and damselflies are both able to produce an aposematic colour signal that is so infrequent in nature. While we redirect the reader to our study in the Journal of Animal Ecology for further detail, we would like to highlight here a result that caught our eye during our investigation: Polythoridae damselflies are probably able to mimic the colour aposematic signal of the glasswing butterflies through a pre-adaptation. Damselfly wings are usually covered with a disorganized matrix of crystalline wax rods, which reflect ultraviolet. Interestingly, these rods were longer and somewhat wider where the white patch is located in Polythoridae damselflies (Fig. 5), even when compared to other closely related species.

The “clear wing complex” has proven a promising study system to investigate questions on multicomponent aposematic signals and their relation to the evolution of imperfect mimicry. Yet, there are still many promising avenues that will require further lab and field research efforts to advance our knowledge in mimicry, one of the most fascinating examples of natural selection in nature. We have confidence that in-depth investigations evaluating the role of multiple characteristics of warning signals will expand across a combination of well-characterized and yet-to-be-explored model systems in the animal ecology community. In this regard, our future work in the “clear wing complex” system aims to further investigate our previous observation that the damselflies are mimicking the flight style of glasswing butterflies. We look forward to offering a comprehensive analysis of further data collected in the field (Fig. 6), quantifying different flight parameters of these animals, aiming to provide a deeper understanding on how flight mimicry has evolved in this system and how it relates to other characteristics of the aposematic signal.

