
This blog post is provided by Fernando Chaguaceda and tells the #StoryBehindThePaper for the article “Short-term apparent mutualism drives responses of aquatic prey to increasing productivity“, which was recently published in Journal of Animal Ecology.
Mobile generalist predators that feed on more than one prey are common in nature, from the majestic whales that spread their feeding habitats from pole to pole to small animals that locally feed on different prey from different habitat patches. Feeding on multiple prey has two important ecological consequences:
The first is that consumers use energy from different pathways (detrital vs. grazing-based, terrestrial vs. aquatic, agriculture-based vs. forest-based, etc.) to build up their own biomass and sustain their populations.
The second is that, in turn, predation pressure on one prey becomes dependent on alternative prey. For example, increasing abundance of one prey can have negative effects on the other prey by sustaining higher predator numbers, leading to Apparent Competition (–,–) (Fig 1A). However, increasing numbers of one prey may also benefit other prey by “diverting the attention” of their shared predator, leading to Apparent Mutualism (+,+) (Fig 1B).
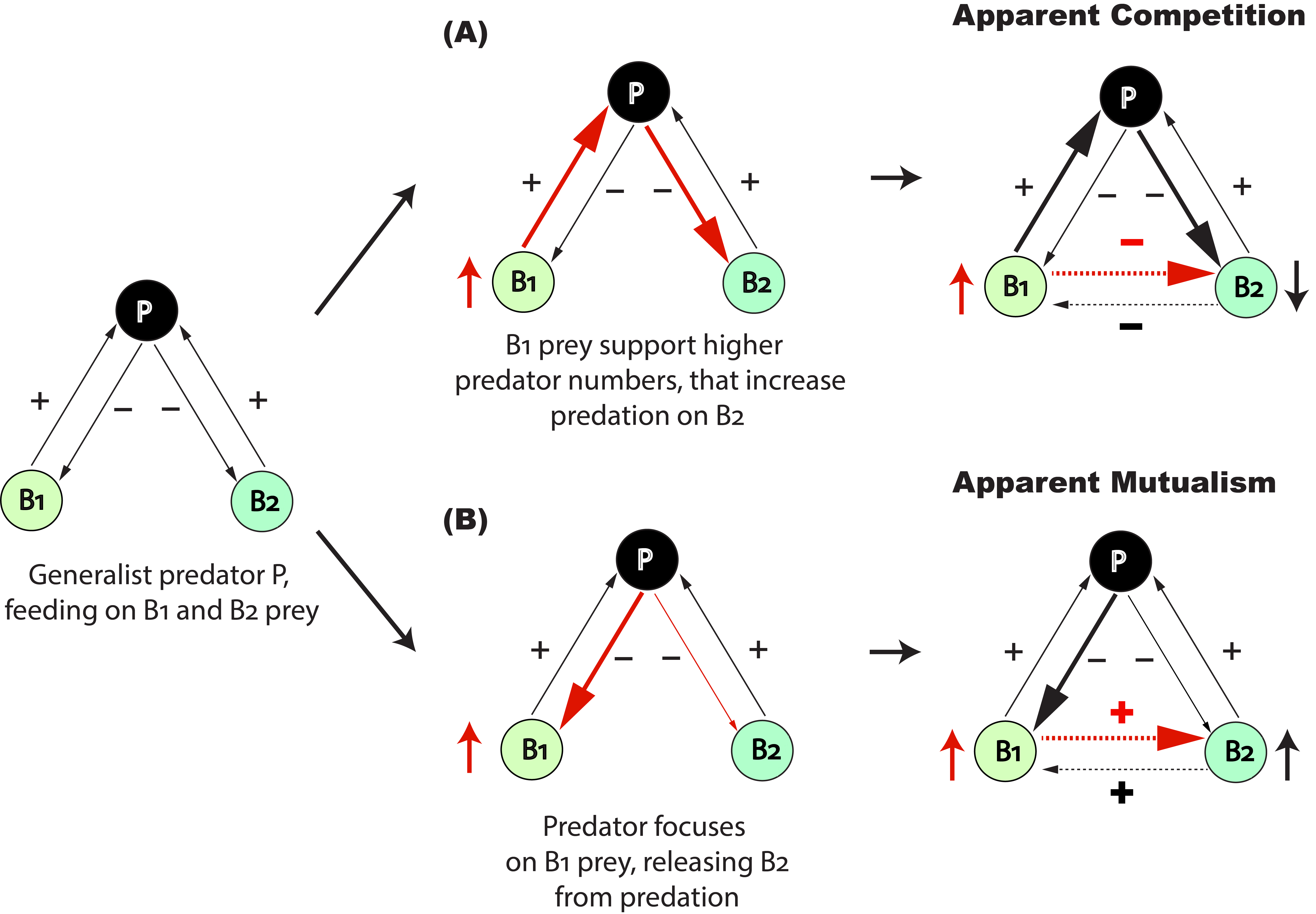
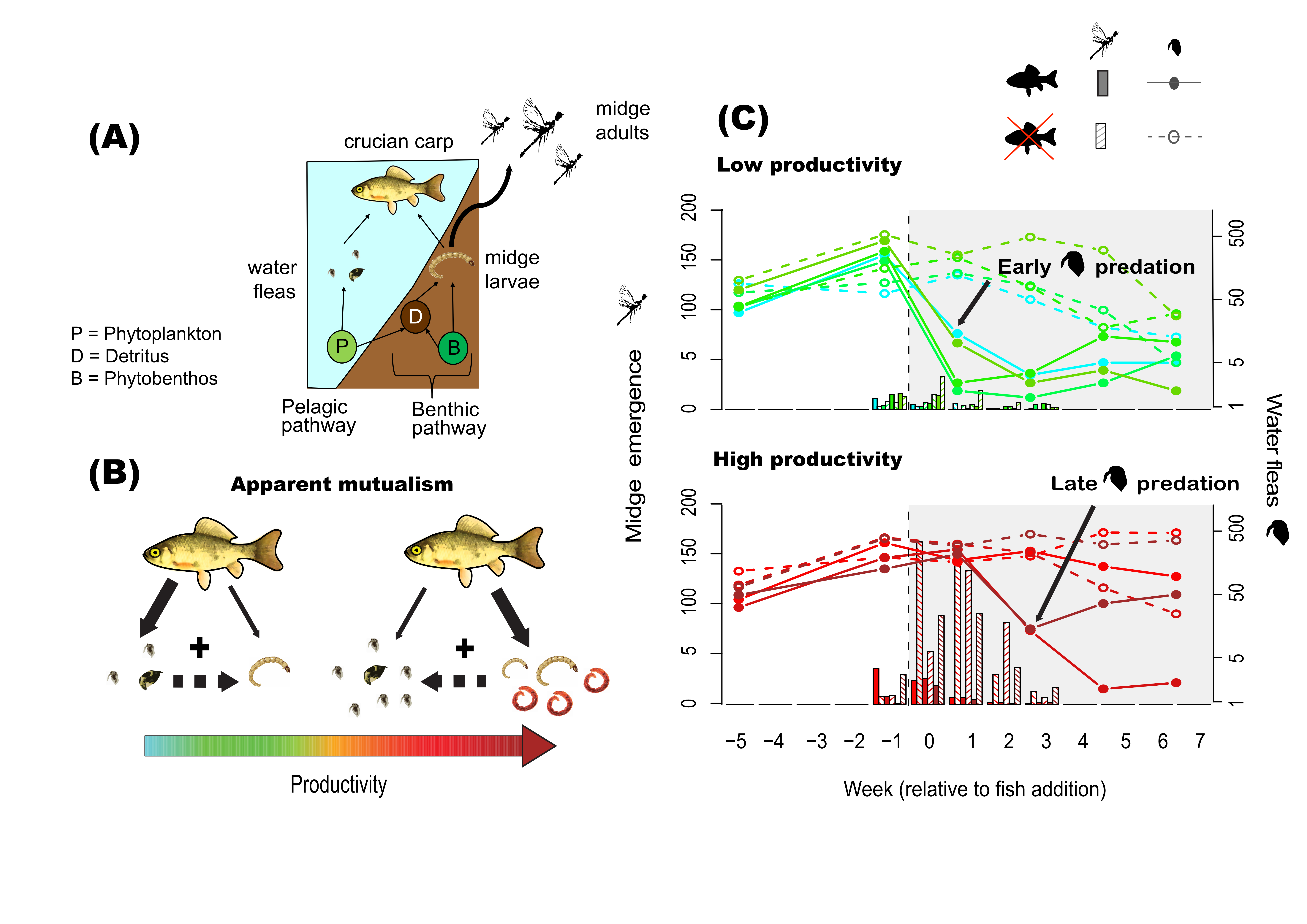
In our study published in the Journal of Animal Ecology, we investigated such indirect prey-prey interactions through shared predation, particularly the mechanisms controlling them over short time frames and in response to increasing productivity. We used a lake food web as a model system, where generalist crucian carps (Carassius carassius) would feed both on water fleas in the water column and midge larvae on sediment surfaces (Fig 2A).
We found that increasing abundance of midges with rising productivity had a positive effect on water fleas abundance, leading to apparent mutualism (+,+) (Fig 2B). By looking at prey depletion trends over time, we were able to link short-term apparent mutualism to the foraging behaviour of the predator, where crucian carps acted as switching predators that shifted from water fleas to midge prey at higher midge abundances. Prey shifting did not only decrease the predation pressure on water fleas at higher productivity, but also delayed it, affecting the dynamics of water flea populations (Fig 2C). Altogether, increasing productivity strongly affected indirect prey-prey interactions through shared predation and its effects on prey community dynamics. Thus, indirect prey-prey interactions can act as the key regulators of community responses to environmental change.
Mesocosm experiments: the product of intense multidisciplinary teamwork
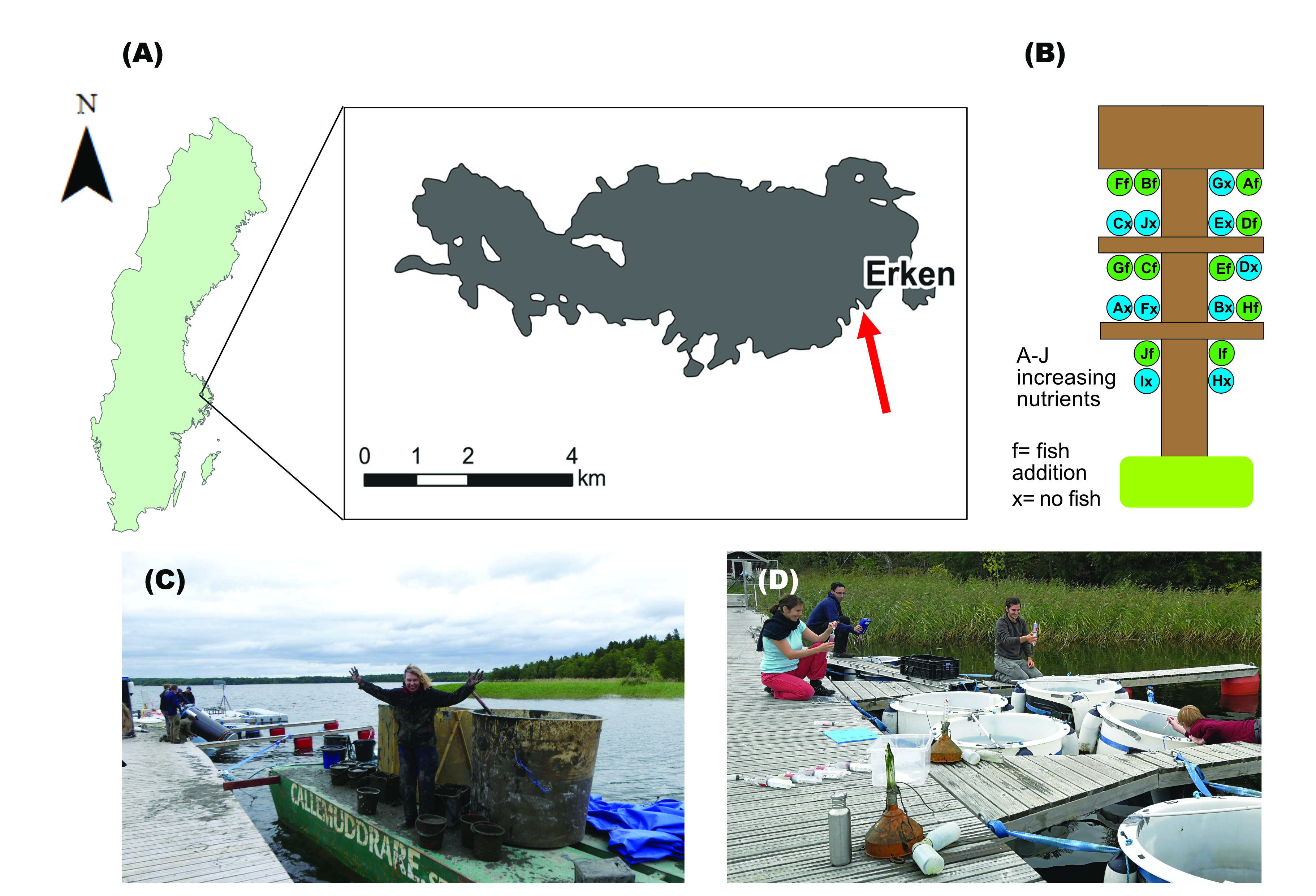
Our study is based on a mesocosm experiment on the shore of Lake Erken at the Erken Laboratory (Fig 3A), which consists of 20 1000L cylinders filled with lake water. We created a nutrient gradient from moderate to highly productive conditions in the presence or absence of crucian carp as a top predator (Fig 3B). The experiment was part of a multidisciplinary project (KAWater), that included > 25 collaborators across several different research fields (e.g. Chemistry, Engineering, Microbiology, Ecology), including many students. Apart from providing information on food-web interactions, this experiment helped us to know more about greenhouse emissions of lakes depending on productivity and food web structure1 , and about fluxes of important fatty acids to terrestrial food webs via midge emergence2.
The experiment started in July 2017 with the enormous effort of the mesocosm deployment (Fig 3C). Professors and students worked together in different teams: some installed periphyton strips, buoys and heavy chains on the mesocosms. Then, others transfered the mesocosms to the dock, while others poured in buckets of sediment leaving a trace of mud (Fig 3C). Finally, 2-3 people would fill the mesocosms with pumped lake water while introducing them in the lake (Fig 3C).

In total, the experiment spanned 14 months. In this time, we experienced seasonal changes at the lake (Fig 4): from cold winter (with 50-cm thick ice!) to the summer blooms of “hairy” Gloetrichia cyanobacteria (Fig 5); from the arrival of birds in the spring to their disappearance behind the fast growing reeds.

Soon after the deployment, algal mats had covered the mesocosms outer walls and abundant schools of fish surrounded the new sheltered habitat. Above the water, only scarecrows kept terns away from breeding (Fig 5). A few weeks later, life had also blossomed in the mesocosms and the effects of the nutrient gradient became noticeable in the benthic algal communities (Fig 6). As soon as midges started emerging in late August, we rushed to catch them with emergence traps. In contrast, we caught the water fleas by filtering the water through a fine mesh. Crucian carps were easy to add into the mesocosm, but extremely difficult to remove. To catch these mistrustful fish, we had to use multiple techniques such as hand-nets, angling and fyke-traps (Fig 6).

Finally, In September 2018, the team gathered again for the mesocosm removal. This time, we relied on heavy machinery (and creativity) by towing the mesocosms by boat so that Björn was able to lift them with his tractor (otherwise designed to pick bundles of straw). Since then, the mesocosms are stored next to the sauna at the Erken laboratory, hoping to be used by another team of researchers.
Looking back, the mesocosm experiment has been a lifetime experience to grow as a scientist and as a person. Thanks to all the ones who gave their hands, their expertise and their creativity during this process (Fig 7). We did an awesome job.

Useful links
- Erken Laboratory: https://www.fieldsites.se/en-GB/research-stations/erken-32652341
- KAWater project on Researchgate: https://www.researchgate.net/project/KAWater-Inland-Water-Ecosystems-in-the-Global-Carbon-Cycle-towards-a-mechanistic-understanding
- Limnology research programme at Uppsala University: https://www.ieg.uu.se/limnology/
Read the paper
Read the full paper here: Chaguaceda, F, Scharnweber, K, Dalman, E, Tranvik, LJ, Eklöv, P. Short‐term apparent mutualism drives responses of aquatic prey to increasing productivity. J Anim Ecol. 2021; 00: 1– 12. https://doi.org/10.1111/1365-2656.13413

Thank you very much for this writing. An interesting investigation.
Muchas gracias, @Campillay-Llanos!