
This blog post is provided by Justin Van Goor, Finn Piatscheck, Derek Houston, and John Nason and tells the #StoryBehindThePaper for their article “Differential effects of nematode infection on pollinating and non-pollinating fig wasps: can shared antagonism provide net benefits to a mutualism?“, which was recently published in the Journal of Animal Ecology.
Justin Van Goor completed his PhD at Iowa State University studying fig nematodes in 2018. He is currently a postdoc studying sperm competition in Caenorhabditis nematodes with Eric Haag at the University of Maryland College Park. Finn Piatscheck was also a PhD student during the data collection for this manuscript and is now a postdoc working with Allen Herre at the Smithsonian Tropical Research Institute. Derek Houston was a postdoc during this data collection and is now the Thornton Chair of Biology at Western Colorado University. John Nason is a professor and Chair of the Department of Ecology Evolution and Organismal Biology at Iowa State University.

Individual species are universally part of broader organismal community networks. The organisms within communities constantly interact with one another via ecologies ranging from obligately mutualistic to strongly antagonistic. These interactions have undoubtedly shaped the evolution of all community associates, but untangling the roles that individual species play on the complicated network with which they interact is very challenging for many systems. Developing knowledge of this complex context would allow for greater insight to how whole communities have co-evolved, and is therefore quite rewarding.
One model system capable of addressing and investigating this context is the fig-fig wasp mutualism. Most of the 750+ fig (Ficus) species worldwide maintain obligate mutualistic associations with a pollinating fig wasp species. Additionally, each fig species is associated with at least one, if not many non-pollinating fig wasps (NPFWs). The ecologies of these NPFWs vary based on species, and can function simply as freeloaders (commensals) or can directly compete with or exploit their host fig and/or pollinating wasp (antagonists). Pollinators, NPFWs, and fig seeds all grow in shared reproductive space (the fig interior), and have tightly synchronous life histories.

Each fig pollinator is also associated with at least one nematode species. Depending on the genus, these nematodes can also range from commensalistic to parasitic. These nematodes share the same reproductive space as pollinators and NPFWs, and latch onto hosts as they are exiting a natal fig. Importantly, pollinators are the only wasp in most fig communities that directly enter a fig and can provide nematodes with nutrition and access to their own reproductive space. Conversely, all Neotropical NPFWs oviposit their eggs from the fig exterior, precluding nematode access to successful reproduction and subsequent dispersal. As such, there should be strong selection against nematode infection of NPFWs in favor of the “proper” pollinating wasp host.
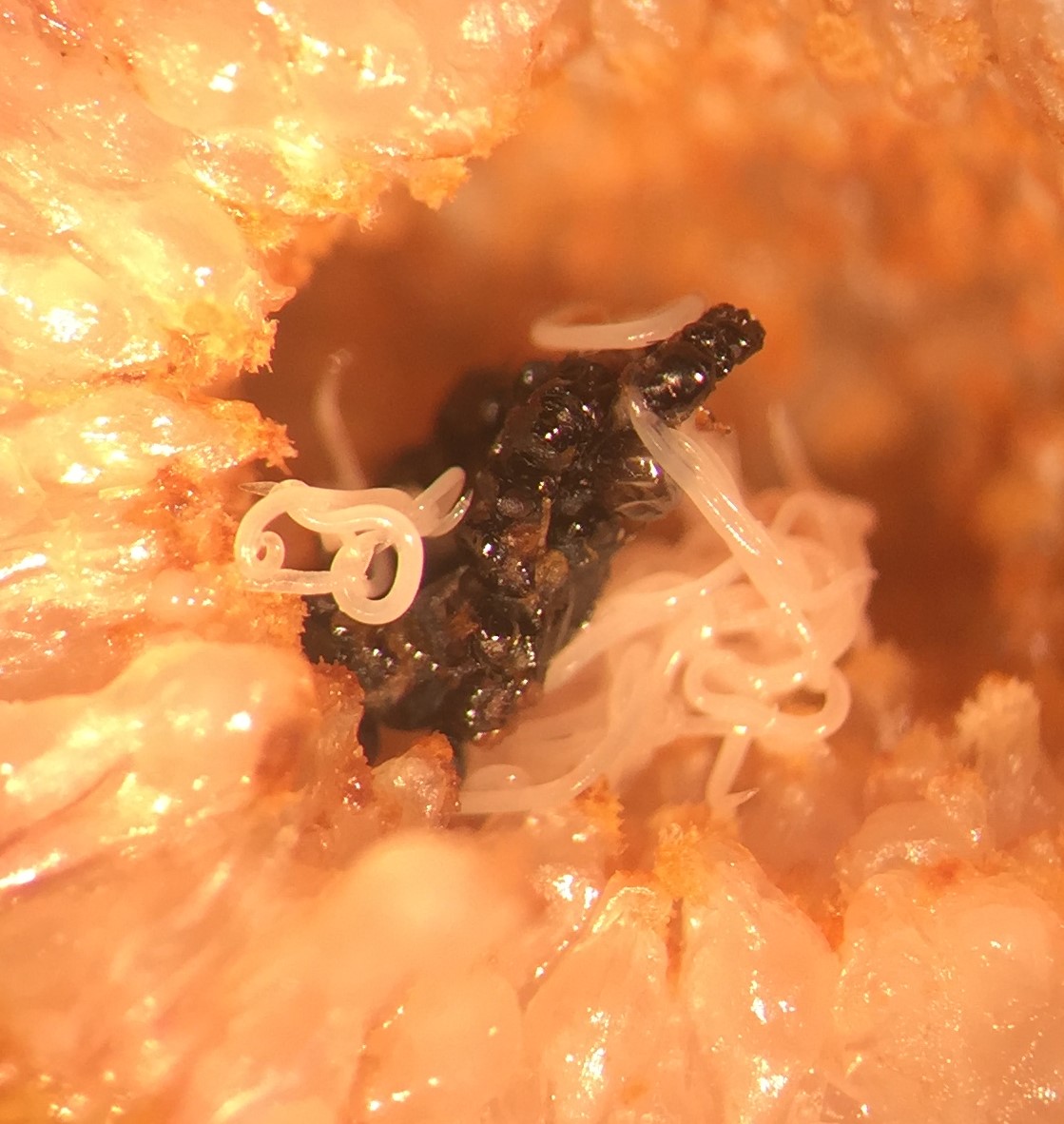
Ficus petiolaris is the only endemic fig species in Mexico’s Baja California Peninsula. Along with its mutualistic Pegoscapus pollinating wasp species, F. petiolaris is associated with at least nine NPFW species. The pollinator is also subject to parasitism by one undescribed Parasitodiplogaster nematode species. The original intent of this work was to describe the effect of nematodeinfection on fig and pollinating fig wasp fitness over geographic space and time. This community context could be applied to better understand how mutualisms evolve in the face of ever-present antagonism.

To address this (and other projects), our lab would drive a pickup truck laden with camping and sampling gear from Iowa to the states of Baja California and Baja California Sur, Mexico, where we repeatedly visited nine GPS-mapped F. petiolaris populations spanning a nearly 750km longitudinal gradient of the peninsula. Collectively, we wound up completing four of these collection trips. When sampling, we would travel to F. petiolaris populations of interest, which were often located on rancher-owned lands hours away from the closest town. We would camp out at these populations for one or two days while sampling, before driving to the nearest town where we would set up a makeshift “laboratory space” in roadside motels for observations/sample processing (much to the amusement/chagrin of the owners/housekeeping staff).

I remember quite vividly the moment that this manuscript “came to life”. It was May 2013, and I was assessing nematode infection in mature, wasp rearing figs using my dissection microscope in our hotel room at the Malarrimo Hotel in Guerrero Negro, BCS (great food, by the way). As wasps were emerging from their natal galls, I watched juvenile, infectious-stage Parasitodiplogaster nematodes performing nictation behavior to contact wasp hosts. Surprisingly, I saw a nematode latch onto an Idarnes flavicollis female wasp. Being a NPFW, I “knew” that this was not supposed to happen, and thus I wrote it off as a fluke (or a very confused nematode). However, afterward I began to see this type of infection occurring more frequently. It seemed to be happening quite regularly, and seemingly to all NPFWs the community had to offer. I even saw juvenile nematodes infecting male NPFWs, which really shouldn’t happen considering males in most species will never leave a natal fig in their lifetime. Infection of any NPFW (male or female) constituted a reproductive dead-end for the nematodes, and yet I was seeing this behavior consistently. Why was this happening? What are the community-level consequences for this? Could nematodes be helping the fig-fig wasp mutualism by infecting antagonistic NPFWs?

To address this, we needed to first assess which NPFWs acted antagonistically with the mutualism. Through our years of collections we were able to amass the wasp contents of over 2400 figs, allowing us to make inferences through correlations. We were then able to determine how frequently wasps were infected by cutting open thousands of tiny (generally <2mm) wasps and counting the numbers of nematodes inside, if infected. By rearing out wasps in “longevity vials” we were able to determine if there was an effect of nematode infection on wasp lifespan, as well as the numbers of nematodes found inside “departing” hosts. While the nematode number in successfully dispersed pollinators is easy to deduce (because they spill out into the fig interior), we needed to be a bit more creative to find out how many nematodes were in successful NPFWs. This involved spending hours standing around receptive figs, aspirators in-hand, hoping to see NPFWs arriving so that we could collect them.

Ultimately, through this work and its direct predecessor (see Van Goor et al 2018); we found that while Parasitodiplogaster infection of F. petiolaris pollinators has a benign effect on wasp (and therefore mutualism) fitness, infection of NPFWs may have drastically stronger effects. Nematode infection appears to significantly limit NPFW dispersal ability, and therefore fitness. Thus, far from being “simple” parasites, nematodes in this system may provide indirect mutualistic benefits to their wasp and fig hosts. Further, this could represent a more ecologically common phenomenon than previously described, where communities are modulated around shared reproductive, energetic, and infectious sources, especially in invertebrate-rich assemblages.

Read the paper
Read the full paper here: Van Goor, J., Piatscheck, F., Houston, D.D. and Nason, J.D. (2021), Differential effects of nematode infection on pollinating and non‐pollinating fig wasps: can shared antagonism provide net benefits to a mutualism?. Journal of Animal Ecology. Accepted Author Manuscript. https://doi.org/10.1111/1365-2656.13495
